Дыхательная функция крови. Кислотно-основное состояние организма презентация
Содержание
- 2. С Т Р О Е Н И Е Г Е М
- 3. Н О Р М А Л Ь Н Ы Е Ф
- 4. ПАТОЛОГИЧЕСКИЕ ФОРМЫ ГЕМОГЛОБИНА HbS – гемоглобин серповидно-клеточной анемии. В
- 5. РЕГУЛЯЦИЯ ПРИСОЕДИНЕНИЯ КИСЛОРОДА К ГЕМОГЛОБИНУ Олигомерная структура гемоглобина обеспечивает быстрое
- 6. КРИВАЯ ДИССОЦИАЦИИ ОКСИГЕМОГЛОБИНА
- 7. ВЛИЯНИЕ рН ВЛИЯНИЕ рН При снижении рН (закислении среды) сродство НЬ
- 8. ДЫХАТЕЛЬНАЯ ФУНКЦИЯ КРОВИ ОСОБЕННОСТИ ОБМЕНА О2 В ЛЕГКИХ и ТКАНЯХ:
- 9. ОСОБЕННОСТИ ОБМЕНА СО2 В ТКАНЯХ и легких В тканях диффундирующий в
- 10. ОСОБЕННОСТИ ОБМЕНА ГАЗОВ
- 11. ГИПОКСИЯ (кислородное голодание)
- 12. КИСЛОТНО-ОСНОВНОЕ СОСТОЯНИЕ Кислотно-основное равновесие представляет собой активность физиологических и физико-химических
- 13. ЗНАЧЕНИЕ ПОСТОЯНСТВА рН ДЛЯ ОРГАНИЗМА РОЛЬ ВНЕКЛЕТОЧНЫХ ИОНОВ ВОДОРОДА оптимальную
- 15. БУФЕРНЫЕ СИСТЕМЫ КРОВИ
- 16. В процессе обмена веществ в организме непрерывно образуются продукты кислотного характера.
- 17. БУФЕРНЫЕ СИСТЕМЫ КРОВИ
- 18. БЕЛКОВАЯ БУФЕРНАЯ СИСТЕМА Второе название – аминокислотная БС. Состоит
- 19. ФОСФАТНАЯ БУФЕРНАЯ СИСТЕМА Фосфатная буферная система составляет около 1-2% от
- 21. ГИДРОГЕНКАРБОНАТНАЯ БУФЕРНАЯ СИСТЕМА Действует как в эритроцитах, так и во всех
- 23. ГЕМОГЛОБИНОВАЯ БУФЕРНАЯ СИСТЕМА Обладает наибольшей мощностью. На него приходится до
- 25. РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ КОС Развитие почечной реакции на смещение
- 27. АММОНИЕГЕНЕЗ АММОНИЕГЕНЕЗ Аммониегенез происходит на протяжении всего почечного канальца, но более
- 28. АЦИДОГЕНЕЗ В процессе ацидогенеза в сутки с мочой выделяется 10-30 ммоль
- 29. ВИДЫ НАРУШЕНИЙ КИСЛОТНО-ОСНОВНОГО СОСТОЯНИЯ Можно выделить следующие причины: 1. Повышенное
- 30. МЕТАБОЛИЧЕСКИЙ АЦИДОЗ ПРИЧИНЫ 1. Повышение содержания кислот в крови Кетоацидоз
- 31. РЕСПИРАТОРНЫЙ АЦИДОЗ ПРИЧИНЫ Причинами являются нарушение вентиляции легких, сопровождающиеся гиповентиляцией:
- 32. МЕТАБОЛИЧЕСКИЙ АЛКАЛОЗ Причины 1. Эндогенный синтез и повышенная секреция в
- 33. СПАСИБО СПАСИБО ЗА ВНИМАНИЕ!
- 34. Скачать презентацию






 сродство НЬ")



")


















Слайды и текст этой презентации
Слайд 1
Описание слайда:
ГЗ «Луганский государственный медицинский университет»
Кафедра медицинской химии
Ассистент Демьяненко Е.В.
Слайд 2
Описание слайда:
С Т Р О Е Н И Е Г Е М О Г Л О Б И Н А
Гемоглобины представляют собой группу родственных белков, содержащихся в эритроцитах.
Гемоглобин выполняет в организме 2 основные функции:
Перенос О2 из легких к периферическим тканям
Перенос СО2 из периферических тканей в альвеолы легких для последующего выведения из организма
Молекулярный кислород плохо растворим в воде (плазме), поэтому практически весь кислород связан с гемоглобином эритроцитов.
Содержание Hb в крови составляет 140-180 г/л у мужчин и 120-160 г/л у женщин.
Гемоглобин - белок, включающий 4 гемсодержащие белковые субъединицы (протомера). Они могут быть представлены различными типами полипептидных цепей: α ("альфа"), β ("бета"), γ ("гамма"), δ ("дельта"), ξ ("кси"). В состав молекулы гемоглобина входят по две цепи двух разных типов.
Гем (железосодержащая часть) соединяется с белковой субъединицей через остаток гистидина координационной связью железа, а также через гидрофобные связи пиррольных колец и гидрофобных аминокислот.
Гем располагается как бы "в кармане" своей цепи и формируется гемсодержащий протомер.
Слайд 3
Описание слайда:
Н О Р М А Л Ь Н Ы Е Ф О Р М Ы Г Е М О Г Л О Б И Н А
Существует несколько нормальных вариантов гемоглобина:
HbР – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, содержится в эмбрионе между 7-12 неделями жизни
HbF – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется после 12 недель внутриутробного развития и является основным после 3 месяцев
HbA – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи. У плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина
HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи
HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочныхвенах его 94-98% от всего количества гемоглобина
HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Слайд 4
Описание слайда:
ПАТОЛОГИЧЕСКИЕ ФОРМЫ ГЕМОГЛОБИНА
HbS – гемоглобин серповидно-клеточной анемии.
В результате точечной мутации в 6-м положении β-цепи вместо аминокислоты глутамата содержится валин. Это приводит к изменению свойств всей молекулы и формирование на поверхности гемоглобина "липкого" участка. При дезоксигенации гемоглобина участок "раскрывается" и связывает одну молекулу гемоглобина S с другими подобными. Результатом является полимеризация гемоглобиновых молекул и образование крупных белковых тяжей, вызывающих деформацию эритроцита и при прохождении капилляров гемолиз.
MetHb (метгемоглобин) - форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного.
Такая форма обычно образуется спонтанно, в этом случае ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления ионов железа в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO (карбоксигемоглобин) - образуется при наличии СО (угарный газ) во вдыхаемом воздухе.
Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни.
HbA1С (гликозилированный гемоглобин) - концентрация его нарастает при хронической гипергликемии и является хорошим скрининговым показателем уровня глюкозы крови за длительный период времени. В норме должен быть не более 7,1%.
Слайд 5
Описание слайда:
РЕГУЛЯЦИЯ ПРИСОЕДИНЕНИЯ КИСЛОРОДА К ГЕМОГЛОБИНУ
Олигомерная структура гемоглобина обеспечивает быстрое его насыщение кислородом в легких и переходом его в оксигемоглобин. Объясняется такой феномен тем, что в легких при присоединении первой молекулы кислорода к железу (за счет 6-й координационной связи) атом железа втягивается в плоскость гема, кислород остается вне плоскости. Это вызывает перемещение участка белковой цепи и изменение конформации первого протомера. Такой измененный протомер влияет на другие субъединицы и облегчает связывание кислорода со второй субъединицей. Это меняет конформацию второй субъединицы, облегчая присоединение последующих молекул кислорода и изменение других протомеров. Четвертая молекула О2 присоединяется в 300 раз легче, чем первая. Взаимовлияние протомеров олигомерного белка друг на друга называется кооперативное взаимодействие. В легких такое
Слайд 6
Описание слайда:
КРИВАЯ ДИССОЦИАЦИИ ОКСИГЕМОГЛОБИНА
Слайд 7
Описание слайда:
ВЛИЯНИЕ рН
ВЛИЯНИЕ рН
При снижении рН (закислении среды) сродство НЬ к О2 снижается и кривая смещается вправо. При повышении рН (защелачивании) увеличивается сродство НЬ к О2 и кривая смещается влево.
Образование большого количества СО2 в тканях способствует увеличению отдачи кислорода за счет снижения сродства НЬ к нему. При выделении СО2 в легких уменьшается рН крови и улучшается оксигенация. CO2 также влияет на диссоциацию НbO2.
ВЛИЯНИЕ ТЕМПЕРАТУРЫ
При снижении температуры отдача О2 оксигемоглобином снижается (сродство повышается, кривая смещается влево), а при ее увеличении ускоряется этот процесс (сродство снижается, кривая смещается вправо).
РОЛЬ 2,3-ДИФОСФОГЛИЦЕРАТА
2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта (пентозо-фосфатного цикла). При снижении концентрации кислорода в эритроцитах повышается содержание 2,3-ДФГ. Он располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями. Функция 2,3-ДФГ заключается в снижении сродства гемоглобина к кислороду.
Это имеет особенное значение при подъеме на высоту, при нехватке кислорода во вдыхаемом воздухе. В этих условиях связывание кислорода с гемоглобином в легких не нарушается, так как концентрация его относительно высока. Однако в тканях за счет 2,3-дифосфоглицерата отдача кислорода возрастает в 2 раза.
НАЛИЧИЕ СОПУТСТВУЮЩИХ ЗАБОЛЕВАНИЙ
Например, при сахарном диабете повышается уровень гликозилированного гемоглобина, который имеет большее сродство к кислороду, чем нормальный гемоглобин. Следовательно, кислород хуже высвобождается из связи с гликозилированным гемоглобином и ткани получают меньше кислорода.
Слайд 8
Описание слайда:
ДЫХАТЕЛЬНАЯ ФУНКЦИЯ КРОВИ
ОСОБЕННОСТИ ОБМЕНА О2 В ЛЕГКИХ и ТКАНЯХ:
Кислород, который поступает в кровь, сначала растворяется в плазме крови.
Кислород, который растворился в плазме крови, по градиенту концентрации проходит через мембрану эритроцита и образует оксигемоглобин (НbО2). Оксигемоглобин - неустойчивое соединение и легко распадается. Прямая реакция называется оксигенацией, а обратный процесс - дезоксигенацией гемоглобина.
Каждая молекула Нb может присоединить 4 молекулы О2, что в пересчете на 1 г Нb означает 1,34 мл О2. Кислородная емкость крови (КЕК) составляет 1,34.
Слайд 9
Описание слайда:
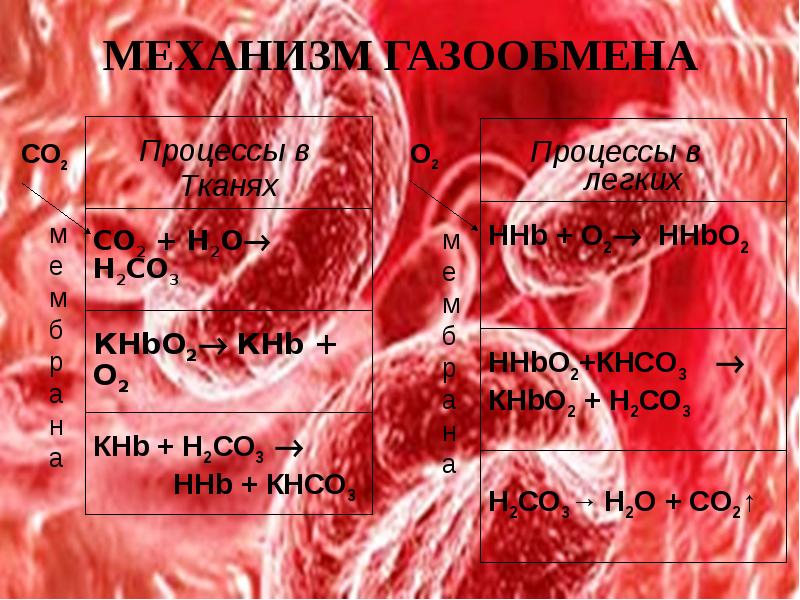
ОСОБЕННОСТИ ОБМЕНА СО2 В ТКАНЯХ и легких
В тканях диффундирующий в кровь из клеток СО2 большей частью (около 90%) попадает в эритроциты. Движущей силой этого процесса является быстрая, постоянно идущая реакция превращения его в угольную кислоту при участии фермента карбоангидразы. Угольная кислота диссоциирует и подкисляет содержимое эритроцита, что улучшает отдачу оксигемоглобином кислорода.
Одновременно с концевыми NH2-группами β-цепей гемоглобина связывается 10-12% карбонат-иона с образованием карбаминогемоглобина (H-HbCO2).
Hb-NH2 + CO2 → Hb-NH-COO– + H+
Остальные бикарбонаты выходят в плазму крови в обмен на ионы хлора (гипохлоремический сдвиг).
В лего чных капиллярах имеется относительно низкая концентрация углекислого газа в альвеолярном воздухе, происходит высокоэффективная диффузия СО2 из плазмы через альвеолярные мембраны и его удаление с выдыхаемым воздухом
уменьшение концентрации СО2 в плазме стимулирует его образование в карбоангидразной реакции внутри эритроцита и снижает здесь концентрацию иона HCO3–,
одновременно высокая концентрация кислорода вытесняет СО2 из комплекса с гемоглобином с образованием оксигемоглобина – более сильной кислоты, чем угольная кислота
диссоциирующие от оксигемоглобина ионы Н+ нейтрализуют поступающий извне ион HCO3– с образованием угольной кислоты. После карбоангидразной реакции образуется СО2, который выводится наружу.
Слайд 10
Описание слайда:
ОСОБЕННОСТИ ОБМЕНА ГАЗОВ
Слайд 11
Описание слайда:
ГИПОКСИЯ
(кислородное голодание)
Слайд 12
Описание слайда:
КИСЛОТНО-ОСНОВНОЕ
СОСТОЯНИЕ
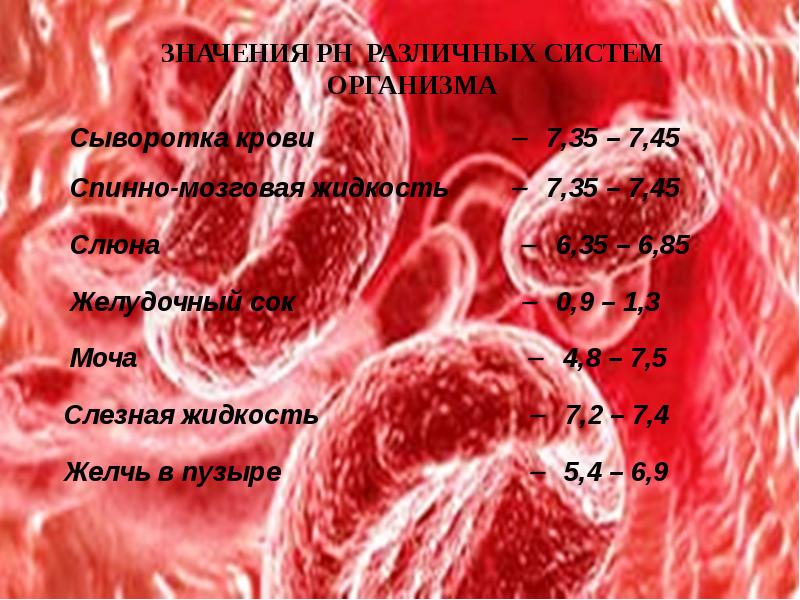
Кислотно-основное равновесие представляет собой активность физиологических и физико-химических процессов, составляющих функционально единую систему стабилизации концентрации ионов Н+. Нормальные величины концентрации ионов Н+ около 40 нмоль/л, что в 106 раз меньше, чем концентрация многих других веществ (глюкоза, липиды, минеральные вещества). Совместимые с жизнью колебания концентрации ионов Н+ располагаются в пределах 16-160 нмоль/л.
Так как реакции обмена веществ часто связаны с окислением и восстановлением молекул, то в этих реакциях обязательно принимают участие соединения, выступающие в качестве акцептора или донора ионов водорода. Роль других соединений – обеспечить неизменность концентрации ионов водорода при жизнедеятельности.
рН внутренних сред организма (крови, лимфы, ликвора, желудочного сока, мочи) оказывает воздействие на жизнедеятельность клеток, тканей, органов и организма в целом.
Значение рН внутренних сред характеризуется значительным постоянством и устойчивостью.
рН – это отрицательный десятичный логарифм концентрации ионов водорода.
рН = - lg [ H+ ]
[ Н+ ] = [ ОН- ] = 10-7 – среда нейтральная – рН = 7
[ Н+ ] > 10-7 – среда кислая – рН < 7
[ Н+ ] < 10-7 – среда щелочная – рН > 7
Слайд 13
Описание слайда:
ЗНАЧЕНИЕ ПОСТОЯНСТВА рН ДЛЯ ОРГАНИЗМА
РОЛЬ ВНЕКЛЕТОЧНЫХ ИОНОВ ВОДОРОДА
оптимальную функциональную активность белков плазмы крови и межклеточного пространства (ферменты, транспортные белки)
Поддержание осмолярности биологических жидкостей
растворимость неорганических и органических молекул
неспецифическую защиту кожного эпителия
отрицательный заряд наружной поверхности мембраны эритроцитов
РОЛЬ ВНУТРИКЛЕТОЧНЫХ ИОНОВ ВОДОРОДА
оптимальной активности ферментов мембран, цитоплазмы и внутриклеточных органелл,
формирования электро-химического градиента мембраны митохондрий на должном
уровне и достаточную наработку АТФ в клетке
Различные заболевания также сопровождаются изменением рН биологических жидкостей. Знание этих закономерностей позволяет прогнозирование нарушения КОС в организме и применять соответствующие методы коррекции.
Слайд 14
Описание слайда:
Слайд 15
Описание слайда:
БУФЕРНЫЕ СИСТЕМЫ КРОВИ
Слайд 16
Описание слайда:
В процессе обмена веществ в организме непрерывно образуются продукты кислотного характера. Конечный продукт питательных веществ – СО2, – накапливается в крови, в результате чего создается непрерывная угроза накопления и повышения концентрации Н+ (уменьшения рН).
В процессе обмена веществ в организме непрерывно образуются продукты кислотного характера. Конечный продукт питательных веществ – СО2, – накапливается в крови, в результате чего создается непрерывная угроза накопления и повышения концентрации Н+ (уменьшения рН).
При нормальном функционировании организма наряду с буферными системами с колебаниями КОС борются физиологические механизмы (почки, печень, кишечник, легкие).
Способность буферных систем поддерживать значение рН не беспредельное. Она определяется буферной емкостью.
Буферная емкость – количество сильной кислоты или щелочи (в ммоль/л), прибавленной к 1 л буфера для смещения рНᵥ раствора на единицу.
Буферная емкость является количественной мерой буферного действия и зависит от концентрации и от соотношения компонентов системы.
Слайд 17
Описание слайда:
БУФЕРНЫЕ СИСТЕМЫ КРОВИ
Слайд 18
Описание слайда:
БЕЛКОВАЯ БУФЕРНАЯ СИСТЕМА
Второе название – аминокислотная БС.
Состоит из белков, большая часть которых представлена альбуминами. Максимальное действие осуществляет в плазме. Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, выполняют роль буфера благодаря своим амфотерным свойствам. В кислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно. В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
МЕХАНИЗМ ДЕЙСТВИЯ
NH3+-CH2-COO-
Слайд 19
Описание слайда:
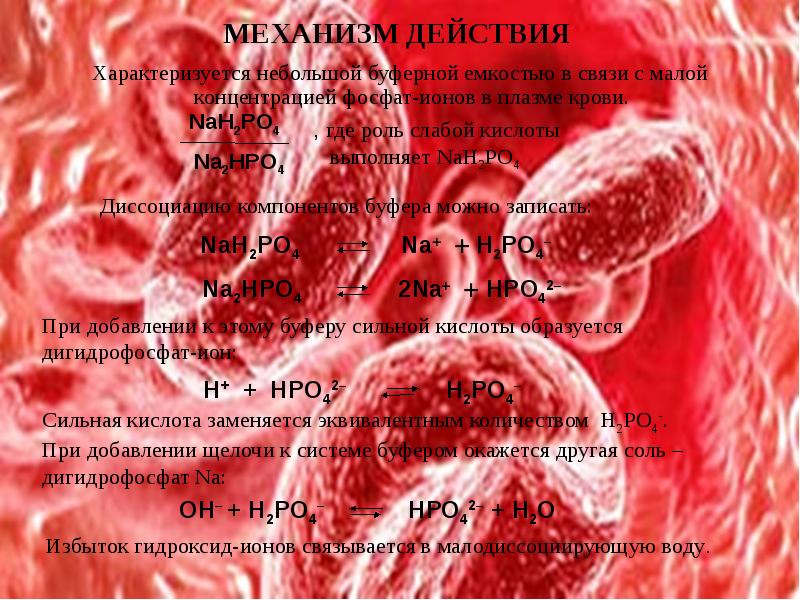
ФОСФАТНАЯ БУФЕРНАЯ СИСТЕМА
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и
до 50% буферной емкости мочи. Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия. Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1.
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи. В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 ↔ NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Слайд 20
Описание слайда:
Слайд 21
Описание слайда:
ГИДРОГЕНКАРБОНАТНАЯ БУФЕРНАЯ СИСТЕМА
Действует как в эритроцитах, так и во всех внеклеточных жидкостях. Это самая мощная внеклеточная система организма.
Состоит из слабой угольной кислоты Н2СО3 (образуется при взаимодействии СО2 с водой под действием фермента карбоангидразы) и натриевой или калиевой соли, образованной этой кислотой и сильным основанием.
H2CO3 или H2CO3
NaHCO3- КHCO3-
СО2 образуется в тканях в результате ферментного окисления углеводов, липидов, белков.
Соотношение гидрогенкарбонат-ионов и угольной кислоты в крови в норме 20:1.
Избыток гидрогенкарбонат-ионов обеспечивает так называемый щелочной резерв крови 25 – 30 ммоль/л химически связанного углекислого газа.
Благодаря работе бикарбонатного буфера концентрация водородных ионов понижается
по двум причинам:
угольная кислота является очень слабой кислотой и плохо диссоциирует
в крови легких благодаря присутствию в эритроцитах фермента карбоангидразы, угольная кислота быстро расщепляется с образованием CO2, удаляемого с выдыхаемым воздухом:
Н2СО3 ↔ Н2О + СО2↑
Слайд 22
Описание слайда:
Слайд 23
Описание слайда:
ГЕМОГЛОБИНОВАЯ БУФЕРНАЯ СИСТЕМА
Обладает наибольшей мощностью. На него приходится до 30% всей буферной емкости крови. В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве (около 8%). Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легк о отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Гемоглобиновой KHb
HHb
Оксигемоглобиновой KHb
HHbО2
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера. Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb – более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Работа гемоглобинового буфера неразрывно связана с дыхательной системой. В легких после удаления СО2 (угольной кислоты) происходит защелачивание крови. При этом присоединение О2 к дезоксигемоглобину H-Hb образует кислоту ННbО2 более сильную, чем угольная. Она отдает свои ионы Н+ в среду, предотвращая повышение рН:
Н-Hb + O2 → [H-HbO2 ] → НbO2 + Н+
В капиллярах тканей постоянное поступление кислот (в том числе и угольной) из клеток приводит к диссоциации оксигемоглобина НbO2 и связыванию ионов Н+ в виде Н-Hb:
НbO2+ Н+→[H-HbO2 ] → Н-Hb + O2
Слайд 24
Описание слайда:
Слайд 25
Описание слайда:
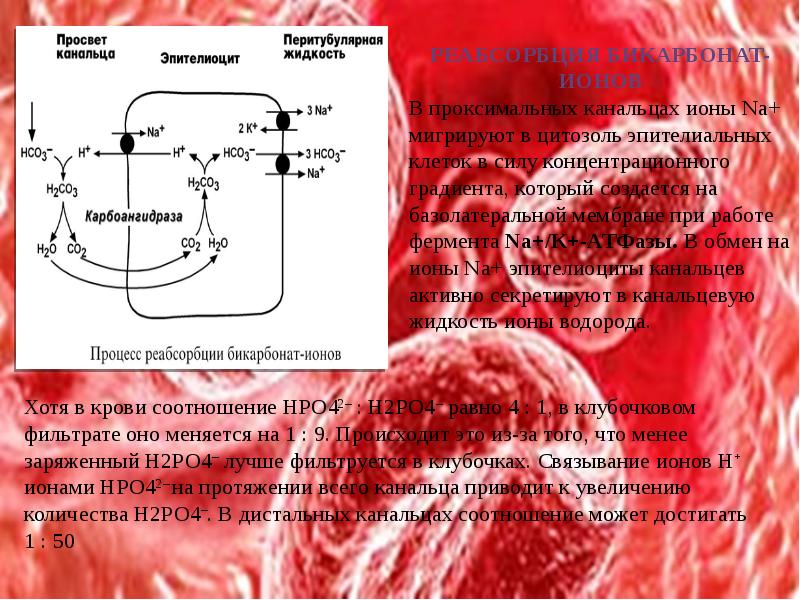
РОЛЬ ПОЧЕК В РЕГУЛЯЦИИ КОС
Развитие почечной реакции на смещение кислотно-основного состояния происходит в течение нескольких часов и даже дней.
Роль почек в регуляции сдвигов КОС заключается в изменении реабсорбции бикарбоната и секреции аммиака и титруемых кислот. Благодаря этим процессам рН мочи постепенно снижается до 4,5-5,2.
Специфические нейрогуморальные механизмы регуляции секреции и реабсорбции ионов Н+ отсутствуют.
В почках активно протекают три процесса, связанных с уборкой кислых эквивалентов:
1. Реабсорбция бикарбонатных ионов HCO3–.
2. Ацидогенез – удаление ионов Н+ с титруемыми кислотами (в основном в составе дигидрофосфатов NaH2PO4).
3. Аммониегенез – удаление ионов Н+ в составе ионов аммония NH4+.
Слайд 26
Описание слайда:
Слайд 27
Описание слайда:
АММОНИЕГЕНЕЗ
АММОНИЕГЕНЕЗ
Аммониегенез происходит на протяжении всего почечного канальца, но более активно идет в дистальных отделах – дистальных канальцах и собирательных трубочках коркового и мозгового слоев. В этих сегментах, в отличие от Na+/H+-антипорта проксимальных отделов, секреция ионов Н+ происходит с участием Н+-АТФазы, локализованной на апикальной мембране эпителиоцита. Ионы HCO3– первичной мочи и секретируемые ионы Н+ образуют угольную кислоту Н2СО3. В гликокаликсе эпителиоцитов фермент карбоангидраза катализирует распад угольной кислоты на СО2 и воду. В результате возникает градиент концентрации углекислого газа между просветом канальцев и цитозолем и СО2 диффундирует в клетки. Внутриклеточная карбоангидраза использует пришедший СО2 и образует угольную кислоту, которая диссоциирует. Ионы НСО3– транспортируются в кровь, ионы Н+ – секретируются в мочу в обмен на ионы Na+. Таким образом, объем реабсорбции НСО3– полностью соответствует секреции ионов Н+. В проксимальных канальцах происходит реабсорбция 90% профильтрованного НСО3–.
Слайд 28
Описание слайда:
АЦИДОГЕНЕЗ
В процессе ацидогенеза в сутки с мочой выделяется 10-30 ммоль кислот, называемых титруемыми кислотами. Фосфаты, являясь одной из этих кислот, играют роль буферной системы в моче. Роль ее состоит экскреции кислых эквивалентов без потерь бикарбонат-ионов за счет дополнительного иона водорода в составе выводимого NaH2PO4 (по сравнению с NaHCO3):
Na2HPO4 + Н2СО3 ↔ NaH2PO4 + NaНСО3
Слайд 29
Описание слайда:
ВИДЫ НАРУШЕНИЙ КИСЛОТНО-ОСНОВНОГО СОСТОЯНИЯ
Можно выделить следующие причины:
1. Повышенное поступление кислых продуктов или недостаточность их удаления.
2. Изменение количества иона НСО3– в сторону увеличения или снижения.
3. Изменение концентрации компонентов буферных систем
Смещение КОС крови в сторону повышения концентрации ионов водорода (снижение рН до 7,0) и уменьшения резервной щелочности ― ацидоз. Смещение КОС крови в сторону понижения концентрации ионов водорода (повышение рН до 7,8) и увеличения резервной щелочности крови ―алкалоз.
Слайд 30
Описание слайда:
МЕТАБОЛИЧЕСКИЙ АЦИДОЗ
ПРИЧИНЫ
1. Повышение содержания кислот в крови
Кетоацидоз (при сахарном диабете, недостатке углеводов в пище при достаточном потреблении белков и жиров, при отравлении алкоголем)
Лактоацидоз (при сепсисе, кровотечении, отеке легких, сердечной недостаточности, при шоке, повышении вязкости крови при остром панкреатите, сахарном диабете, лейкемии, хроническом алкоголизме).
2. Потеря бикарбонатов
С кишечным, панкреатическим и билиарным секретами при диареях и фистулах кишечника и желчного пузыря, дренировании поджелудочной железы.
3. Недостаточное выведение ионов Н+ почками
При уменьшении числа функционирующих нефронов при хронической почечной недостаточности или поражение канальцев.
Слайд 31
Описание слайда:
РЕСПИРАТОРНЫЙ АЦИДОЗ
ПРИЧИНЫ
Причинами являются нарушение вентиляции легких, сопровождающиеся гиповентиляцией:
Повреждения или заболевания легких (пневмония, фиброз, отек легких)
Все случаи механической асфиксии
Повреждения или заболевания дыхательных мышц (нехватка калия, боли после операции, травмы, накопление жировых отложений)
Угнетение дыхательного центра (опиаты, барбитураты), неправильный режим ИВЛ
Бронхиальная астма, эмфизема, бронхит.
При недостаточной вентиляции легких рСО2 способен достичь 140-150 мм рт.ст.
Слайд 32
Описание слайда:
МЕТАБОЛИЧЕСКИЙ АЛКАЛОЗ
Причины
1. Эндогенный синтез и повышенная секреция в кровь ионов НСО3–:
обкладочными клетками желудка при неукротимой рвоте, фистуле желудка, кишечной непроходимости и тд.
РЕСПИРАТОРНЫЙ АЛКАЛОЗ
Причины
1. Возбуждение дыхательного центра.
изолированный ацидоз церебральной жидкости, который является остаточным явлением после компенсированного ацидоза крови.
субарахноидальное кровотечение, при этом происходит стимуляция дыхательного центра продуктами гемолиза
при циррозе печени и сепсисе происходит стимуляция дыхательного центра токсинами и циркулирующими метаболитами.
2. Лихорадочные состояния.
3. Неправильный режим искусственной вентиляции легких.
Слайд 33
Описание слайда:
СПАСИБО
СПАСИБО
ЗА
ВНИМАНИЕ!
Скачать презентацию на тему Дыхательная функция крови. Кислотно-основное состояние организма можно ниже: